January 15, 2020 feature
Shrinking dinosaurs and the evolution of endothermy in birds
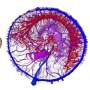
 restored as a shoreline-dwelling predator and scavenger. Artwork by Bob Nicholls (paleocreations.com). Credit: Natural History Museum, London. Credit: Science Advances, doi: 10.1126/sciadv.aaw4486")
The evolution of endothermy (thermoregulation by metabolic means) represents a major transition in vertebrate history. However, the process of endothermy evolution and its timeline in birds and mammals remains controversial. In a new report on Science Advances, Enrico L. Rezende and a team of researchers at the Center of Applied Ecology and Sustainability, and the Institute of Environmental and Evolutionary Sciences in Chile, combined a heat transfer model with theropod body size data. The researchers then reconstructed the evolution of metabolic rates along the bird stem lineage. The results suggested that a reduction in size constituted the path of least resistance for endothermy to evolve—maximizing thermal niche expansion, while reducing costs of elevated energy requirements.
As a result, the researchers hypothesize that metabolism would have increased with miniaturization during the Early-Middle Jurassic period (approximately 180 to 170 million years ago) to result in a gradient of metabolic levels in the theropod phylogeny. While basal theropods may have exhibited lower metabolism rates, the more recent non-avian lineages were likely decent thermoregulators with improved metabolism. The analysis provided a tentative sequence in time of the key evolutionary transitions, for the emergence of small, endothermic and flying feathered dinosaurs.
The evolution of endothermy in birds and mammals is an important transition during vertebrate evolution providing an extraordinary instance in evolutionary convergence between groups, pivotal to their widespread geographic distribution and ecological success. Although several groups of invertebrates and vertebrates can raise their temperatures above ambient, the ability to maintain high and constant body temperature via endogenous heat production at rest is exclusive to birds and mammals alone. The observation explained their greater mobility, stamina and tolerance across a range of conditions. However, this strategy is energetically costly and has no virtual trace in the fossil record, therefore the pace and mode of endothermy remains controversial during vertebrate evolution.
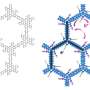
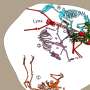
 The cost-benefit to switch from ectothermy to endothermy for different ranges of body size was quantified with the Scholander-Irving model, which describes how a rise in metabolism at rest (cost) increases the thermal niche Tb − Ta (benefit). Because there is no thermal gradient between the organism and the environment in the absence of heat production, this curve intersects the abscissa at Tb = Ta when MR = 0 (8). The solid blue and red lines depict the metabolic curves of a typical ectotherm and endotherm, respectively, and the open symbols depict the maximal thermal gradient Tb − Ta possible with resting metabolic rates, used in our model (Eq. 2). (B) A reduction in body size, consistent with the one described from ancestral theropods to basal birds (22), constitutes the evolutionary path of least resistance as the energy costs of being large are traded for those of being endothermic. Credit: Science Advances, doi: 10.1126/sciadv.aaw4486")
To understand the origin and reason for endothermy during the evolution of birds and mammals, the scientists considered two fundamental questions. What are the costs and benefits of this strategy compared to ectothermy? And what conditions favored a transition toward endothermy? Rezende et al. addressed the questions using the Scholander-Irving model of heat transfer, used to study thermoregulation in endotherms for more than 60 years. The relationship was rarely used for ectotherms, but since all living organisms produced endogenous heat, the model remained applicable under thermal steady state. They made this crucial assumption to avoid using complex dynamic models that are often applied to ectothermy, which could render the present analyses difficult.
They quantified the costs of endothermy as mass-independent energy expenditure, where the benefits included greater mobility and foraging efficiency, predator avoidance, tolerance to and colonization of a wide range of environmental conditions, with increased growth rates and homeostasis. Rezende et al. quantified the thermal niche that organisms could occupy and its expansion to estimate the net benefit of endothermy. They then calculated the cost-benefit of adopting an endothermic lifestyle with reference to the ectothermic ancestor and endothermic descendent. As originally proposed by the evolutionary biologist Bran K. McNab, when the scientists replicated these calculations with exact body size estimates, the results showed that smaller sizes reduced the energy costs to evolve towards endothermy.
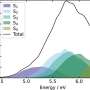
 Theropod phylogeny with branches color-coded according to reconstructed metabolic levels. (B) Scaling of metabolic rate versus body mass for ectotherms (MR = 0.68mass0.75) and endotherms (MR = 3.4mass0.75) and the predicted trajectory of the bird stem lineage during the transition from ectothermy to endothermy. Dashed lines show fold differences between ectotherms and endotherms (1× to 5×); open and closed symbols depict reconstructed values for the bird stem lineage and the tips of the phylogeny, respectively. (C) Scaling of thermal conductance C and body mass for ectotherms (C = 2.5mass0.5) and endotherms (C = 1.0mass0.5), fold differences from 2.5× to 1×. (D) Thermal gradient and fold differences calculated with Eq. 1 (in the publication) and values in (B) and (C). The log-log linear trajectories connecting MR and C of the ectothermic ancestor and the endothermic descendant, as well as the resulting trajectory in thermal gradient, are shown with the continuous lines. Credit: Science Advances, doi: 10.1126/sciadv.aaw4486")
The team explored how this heat transfer model combined with phylogenies and body size reconstructions, shed light on the evolution of endothermy in birds and their theropod ancestors. Rezende et al. estimated the costs of evolving endothermy along the bird stem lineage using reconstructed ancestral body sizes, based on the fossil record. To quantify the energy costs within alternative scenarios they simulated the evolution of body size along the lineage and obtained the distribution of cost per degree in the model. They assumed an undirected Ornstein-Uhlenbeck (OU) model between a mass of 10 g and 100,000 kg with a mean evolutionary rate equivalent to those reported for theropods. The simulations indicated a marked decrease in energy costs per degree with miniaturization. The scientists explained the reduced costs using two phenomena.
First, the expansion in thermal niche based on an increase in metabolic rate (MR) was disproportionately higher in larger ectotherms due to their capacity to maintain a high body temperature (Tb). This was accompanied with a relatively low mass-independent metabolic rate, due to initial homeothermy (stable thermoregulation). Accordingly, they noted that larger the starting size of the ectothermic ancestor, cheaper the transition to endothermy. Second, during miniaturization, the animals traded the energy costs of being large for being endothermic. The results explained how the high energy turnover rates evolved, regardless of their impact on food and water requirements. Despite inherent variation in resource availability in evolutionary time—smaller sizes and higher energy turnover rates were systematically favored in this lineage. Outcomes of the work qualitatively agreed with other models on the evolution of endothermy.
 The miniaturization from Tetanurae to basal birds inferred from the fossil record, contrasted against 100 simulated size trajectories starting from the same ancestral body size for illustrative purposes (note that for the subsequent full null model, the ancestral body size is allowed to vary). The error represents the SD in reconstructed values across 20 candidate trees. (B) The frequency distribution of body mass ratios obtained across 10,000 simulated body size trajectories (histogram) and the energy costs to evolve endothermy expressed per degree Celsius (Eq. 2 in the publication) under this null model (gray symbols). In this case, the ancestral body size was obtained from a uniform distribution ranging between 10 g and 100,000 kg. The empirical estimate in the bird stem lineage is shown in red. The region in which a reduction in body size would compensate for the energy costs of evolving endothermy, enabling the population to increase in a scenario of constant resources, is highlighted in gray. The arrow depicts the expected population fold increase, given the observed body size reduction in the bird stem lineage as endothermy evolved. These analyses indicate that the energy costs to evolve endothermy are reduced with miniaturization and, as a result, population size may have increased despite the metabolic costs of an endothermic lifestyle. Credit: Science Advances, doi: 10.1126/sciadv.aaw4486")
The size reduction in bird stem lineage, closely matched the theoretical path of least resistance for endothermy to evolve, then Rezende et al. reconstructed how the phenomenon might have unfolded in the therapod phylogeny. The process indicated the rise of MR (metabolic rate) spanning most of the Early-Middle Jurassic (approximately 180 to 170 million years ago), involving therapod groups where the occurrence of protofeathers and feathers were already ubiquitous. They also suggested highly diverse metabolic rates across the concurrent lineages of Coelurosauria, Maniraptora and Paraves, which could account for the emergence and diversification of these groups during the Late Jurassic period. The work could also explain the abnormally high diversity of Coelurosauria at intermediate body sizes (30 to 300 kg) compared to other dinosaur groups. The research team further demonstrated a niche-fulling model of adaptive radiation in Mesozoic dinosaurs with exceptional rates of body size reduction in the bird stem lineage (i.e. within basal nodes of Coelurosauria and Paraves), however, they did not propose a suitable evolutionary hypothesis to account for the outcome.
In this way, Enrico L. Rezende and colleagues observed two exceptional phenomena during the evolution of birds; (1) a sustained (but not necessarily gradual) miniaturization spanning millions of years and (2) the emergence of endothermy. They used reconstructions to suggest concomitant evolution of endothermy with a decrease in size along the bird stem lineage, as also previously proposed for mammals. The corresponding therapod clades exhibited a whole spectrum of MR. The interpretations indicate that endothermy preceded the evolution of flight and the marked reduction in body size during the evolution of bird stem lineage was accompanied by a major shift in metabolic levels.
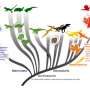
 Reconstructed temporal course of metabolic evolution in the bird stem lineage, with dashed lines showing how reconstructions change assuming that either Paraves or Neornithes were fully endothermic instead of the basal bird [for calculations with Neornithes, we assumed a body size of 150 g based on estimates for Vegavis and a time estimate of 100 Ma ago]. The fold increase in MR was calculated by dividing the reconstructed MR during the transition to endothermy by the MR expected for a similar-sized ectotherm and is therefore dimensionless and independent of body size. (B) The evolutionary path of least resistance from ectothermy to endothermy includes inertial homeothermy as a transitional stage, followed by an increase in metabolism concomitantly with a reduction in size. (C) Hypothetical sequence of evolutionary transitions in the bird stem lineage, which combines results from this study with phylogenetic reconstructions of epidermal structures and capacity for active flight (see the main text). Credit: Science Advances, doi: 10.1126/sciadv.aaw4486")
The proposed scenario explained why theropods systematically favored an expensive lifestyle (higher energy turnover rate) despite its energy costs, and sustained miniaturization preceding the origin of birds. Furthermore, a state of inertial homeothermy constituted a necessary transitional state with low metabolic costs. If large ancestral theropods were physiologically committed to homeothermy, Rezende et al. then expect them to have departed from ectothermic metabolic allometry (relationship of body size to shape, anatomy, physiology and behavior) with miniaturization. During evolutionary time, it is reasonable to expect lineages to exploit newly opened niches and eventually diversify. The outcomes of this work are preliminary; they simply highlight that there is likely more to the evolution of endothermy in archosaurs, dinosaurs and present-day birds. The results form a working hypothesis that can in future studies be combined with previous analyses on the evolution of size and other characteristics in bird stem lineage to form well-defined interpretations of temporal sequences during key evolutionary transitions.
More information: Enrico L. Rezende et al. Shrinking dinosaurs and the evolution of endothermy in birds, Science Advances (2020). DOI: 10.1126/sciadv.aaw4486
D. K. Zelenitsky et al. Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins, Science (2012). DOI: 10.1126/science.1225376
McNab, Brian K. "The Evolution of Endothermy in the Phylogeny of Mammals." The American Naturalist, vol. 112, no. 983, 1978, pp. 1–21. JSTOR, www.jstor.org/stable/2460134. Accessed 15 Jan. 2020.
Journal information: Science Advances , Science , American Naturalist
© 2020 Science X Network